|
Wie DNA-Reparaturprozesse die Darmkrebsentstehung beeinflussen
Wie DNA-Reparaturprozesse die Darmkrebsentstehung beeinflussen
Der Darm ist das größte Organ des Menschen und hat vielfältige Funktionen, die von der Nahrungsaufnahme bis hin zur Immunabwehr reichen. Die Ernährung ist ein wichtiger Faktor für die Darmgesundheit und spielt eine wesentliche Rolle bei der Entstehung von Darmkrebs, der dritthäufigsten Krebserkrankung weltweit. Krebserzeugende Nahrungsbestandteile können das Erbgut schädigen und so Krebs auslösen. Dagegen schützen uns DNA-Reparaturprozesse, die durch Naturstoffe moduliert werden können.
Gefährliche Nahrung

In unserer Nahrung finden sich zahlreiche Stoffgruppen, die mit der Entstehung von Darmkrebs verknüpft sind (Abb.1). Diese können bei der Zubereitung der Lebensmittel erzeugt werden (z.B. beim Braten, Grillen und Räuchern) oder bereits in der Nahrung vorhanden sein. Neuere Studien weisen darauf hin, dass vor allem N-Nitrosoverbindungen in Kombination mit Häm-Eisen eine herausragende Bedeutung bei der Entstehung von Darmkrebs zukommt [1]. Zu den N-Nitrosoverbindungen (NOC) zählen Nitrosamide und Nitrosamine, die überwiegend in Lebensmitteln zu finden sind, aber auch in Kosmetika und im Tabakrauch vorhanden sind. Darüber hinaus können diese Stoffe im Magen und Dickdarm erzeugt werden. NOC besitzen alkylierende Wirkung und schädigen die DNA, wobei mutagene und kanzerogene DNA-Basen wie N7-Methylguanin (N7-MeG) und O6-Methylguanin (O6-MeG) gebildet werden.
Abb.1 An der Darmkrebsentstehung beteiligte karzinogene Stoffe. A 2-Amino-1-methyl-6-phenylimidazo(4,5-b)pyridin (PhIP) ist ein heterozyklisches aromatisches Amin, das bei der Nahrungszubereitung entsteht. B N-Nitrosodimethylamin gehört zur Klasse der N-Nitrosoverbindungen, die in Lebensmitteln vorkommen und endogen im Magen-Darm-Trakt gebildet werden.
C Häm-Eisen ist vor allem in rotem Fleisch wie z.B. Rindfleisch zu finden und spielt eine physiologische Rolle im Sauerstofftransport.
Häm enthält ein zentrales Eisen(II)-Ion und ist die prosthetische Gruppe von Hämoglobin sowie Myoglobin, die beide eine zentrale Rolle im Sauerstofftransport spielen. Häm-Eisen kommt in höheren Mengen in rotem Fleisch (Rindfleisch) vor, dessen Verzehr mit einem signifikant erhöhten Risiko für Darmkrebs verknüpft ist [2]. Als zugrunde liegende Mechanismen der Hämassoziierten Darmkanzerogenese werden unter anderem eine gesteigerte Zellproliferation und die Induktion von prä-mutagenen DNA-Schäden im Dickdarm diskutiert. Häm-Eisen katalysiert zum einen die Bildung von reaktiven Sauerstoffspezies, die DNA-Schäden wie 8-Oxoguanin (8-OxoG) verursachen. Als Sekundärprodukte entstehen dabei reaktive Aldehyde wie Malondialdehyd und 4-Hydroxy-nonenal, welche ebenfalls DNA-Addukte bilden. Zum anderen zeigten mehrere Studien, dass Häm-Eisen zu einer verstärkten endogenen Bildung von NOC führt, die wiederum DNA-Läsionen wie O6-MeG induzieren.
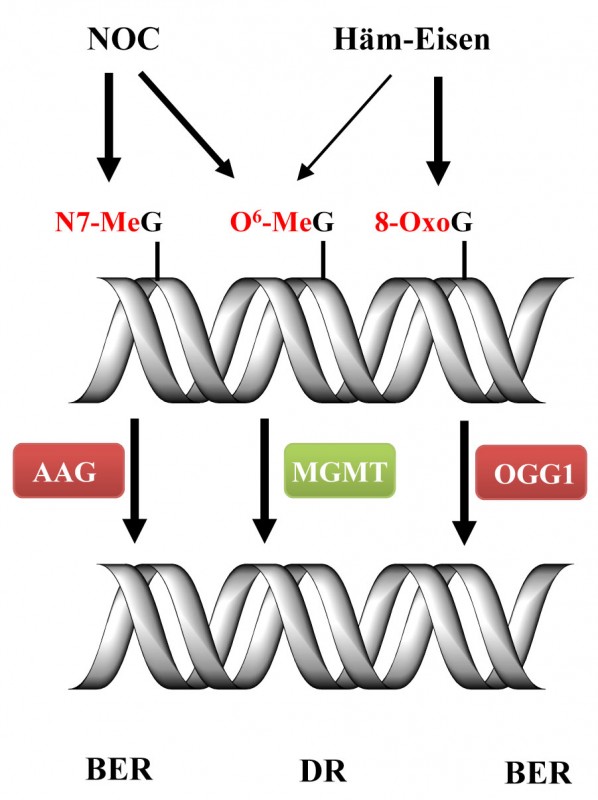
Verschiedene DNA-Reparaturprozesse sind an der Behebung dieser kritischen Schäden beteiligt. Das DNA-Reparaturprotein O6-Methylguanin-DNA-Methyltransferase (MGMT) ist für die Reparatur von O6-MeG zuständig, wohingegen die Basenschäden 8-OxoG und N7-MeG im Rahmen der Basenexzisionsreparatur (BER) durch spezifische DNA-Glykosylasen entfernt werden (Abb.2).
Abb.2 Reparatur von DNA-Schäden, induziert durch Häm-Eisen und NOC. NOC verursachen vor allem N7-Methylguanin (N7-MeG)-DNA-Addukte, die durch das Reparaturprotein N-Alkyladenin-DNA-Glykosylase (AAG) im Rahmen der Basenexzisionsreparatur (BER) behoben werden. Sowohl NOC als auch Häm-Eisen generieren O6-Methylguanin(O6-MeG)-DNA-Addukte, die der direkten Reparatur (DR) mittels MGMT unterliegen. Weiterhin induziert Häm-Eisen die DNA-Läsion 8-Oxoguanin (8-OxoG), die durch die 8-Oxoguanin-DNA-Glykosylase 1 (OGG1) bei der BER entfernt werden.
Eine wichtige Barriere: die DNA-Reparatur
Um die Bedeutung von DNA-Reparaturfaktoren bei der Darmkrebsentstehung genauer zu untersuchen, haben wir ein Mausmodell der kolorektalen Kanzerogenese etabliert (Abb.3a). Dieses beruht auf der alkylierenden Verbindung Azoxymethan (AOM), die mit den NOC verwandt ist und als Tumorinitiator wirkt. Kombiniert wird AOM mit dem negativ geladenen Polysaccharid Dextran-Natriumsulfat (DSS), das eine Entzündung des Dickdarms (Colitis) auslöst und so als Tumorpromotor wirkt. Mithilfe eines schonenden Miniendoskopieverfahrens sind wir in der Lage, die Tumoranzahl im Dickdarm zu quantifizieren und das Tumorwachstum über die Zeit zu verfolgen (Abb.3b). Die durch AOM/DSS erzeugten Dickdarmtumore in Mäusen weisen typische Eigenschaften humaner Tumore auf, wie z.B. Mutationen im KRAS-Onkogen, eine veränderte subzelluläre Lokalisation von ß-Catenin und eine verstärkte Expression des Inflammationsmarkers COX-2 (Abb.3c). Unsere Studien mit DNA-Reparaturkompetenten Mäusen und transgenen Mausstämmen mit Defekten in relevanten DNA-Reparaturgenen (MGMT-/-, AAG-/-) zeigen, dass sowohl MGMT als auch AAG gegenüber der NOC-induzierten kolorektalen Kanzerogenese schützen (Abb. 3d) [1, 3]. In Einklang hierzu wurde beobachtet, dass MGMT bei rund 40% aller sporadischen Dickdarmkarzinome des Menschen bereits im frühen Stadium inaktiviert und mit Mutationen im KRAS-Onkogen assoziiert ist.
Zurzeit wird von uns die Bedeutung des DNA-Reparaturproteins Poly(ADP-Ribose) Polymerase-1 (PARP-1) bei der Colitis-assoziierten Dickdarmkrebsentstehung eingehender untersucht. Diese synthetisiert nach Aktivierung durch DNA-Strangbrüche das Biopolymer Poly(ADP-Ribose) [4] und reguliert so unter anderem die Basenexzisionsreparatur, aber auch entzündliche Prozesse [5].
Abb.3 AOM/DSS-Modell der kolorektalen Kanzerogenese. A Schema des AOM/DSS-Modells. B Miniendoskopische Aufnahmen von gesunder Darmschleimhaut (links) und einem großen Dickdarmtumor (rechts). C Immunhistochemische Färbung von ß-Catenin (grün) und COX-2 (violett) im Tumorgewebe. Blau dargestellt sind die Zellkerne. D Tumoranzahl in DNA-Reparatur-kompetenten, wildtypischen Mäusen (WT) sowie in DNA-Reparaturdefekten Mäusen (MGMT-/- und AAG-/-) 8 Wochen nach Injektion von 5?mg AOM /kg Körpergewicht (n? 10). **, p<0,001 im Vergleich zum WT.
Modulation der DNA-Reparatur durch Naturstoffe
DNA-Reparaturproteine wie MGMT und PARP-1 enthalten kritische Thiol-Gruppen, die von essenzieller Bedeutung für deren Aktivität sind. Bei der Reparatur von O6-MeG-DNA-Addukten wird die Methylgruppe von der geschädigten DNA-Base auf die Thiol-Gruppe des katalytisch aktiven Cystein-Rests von MGMT übertragen. Eine Oxidation oder kovalente Modifikation des Cys-145 führt zum Verlust der Reparaturaktivität. Bisher sind einige Stoffe bekannt, welche die Aktivität von MGMT auf Protein- und/oder Aktivitätsebene modulieren. Antioxidantien wie N-Acetylcystein und Naturstoffe wie Curcumin können die Aktivität von MGMT erhöhen [6], wohingegen das Polyphenol Resveratrol die Expression von MGMT herunterreguliert [7]. Die natürlich vorkommende Disulfidverbindung ?-Liponsäure (LA) besitzt ebenfalls antioxidative Eigenschaften und steigert den zellulären Vorrat an Cystein und Glutathion [8], die wichtige Komponenten für die MGMT-Synthese bzw. Aktivität darstellen. LA weist eine aktivierte Disulfidfunktion auf, weshalb sie potenziell mit anderen Thiol-Gruppen abreagieren kann.
Um den Einfluss von LA auf MGMT genauer zu untersuchen, haben wir zunächst das Reparaturprotein rekombinant exprimiert. Es konnte gezeigt werden, dass LA die Aktivität von MGMT in vitro durch Modifikation des katalytischen Cys-145 inhibiert, wohingegen deren reduzierte Form Dihydroliponsäure keinen Einfluss hatte [9]. Die Inkubation von Dickdarmkrebszellen mit LA führte zur Abnahme der zellulären MGMT-Aktivität und nachfolgend zur Depletion von MGMT auf Proteinebene, ohne die MGMT-mRNA-Expression zu beeinflussen (Abb.4).
Gleichzeitig bewirkte LA eine Autophagieinduktion, wobei diese nicht ursächlich mit dem MGMT-Abbau verknüpft war. Als Folge der MGMT-Degradation sind die LA-behandelten Zellen empfindlicher für die DNA-schädigende Wirkung von SN1-Alkylantien wie dem Zytostatikum Temozolomid (TMZ), das eine deutlich größere Menge an O6-MeG-DNA-Addukten erzeugen kann (Abb.4). Zudem erhöhte LA die Zytotoxizität von TMZ in TMZ-resistenten Dickdarmkrebszelllinien, sodass LA möglicherweise unterstützend in der Tumortherapie eingesetzt werden könnte.
Abb.4 Zelluläre Wirkung von ?-Liponsäure auf DNA-Reparatur und Zelltod. Erläuterungen sind im Text gegeben.
Einleitung des Zelltods durch Naturstoffe
Mittlerweile verdichten sich die Hinweise, dass LA neben ihren antioxidativen Eigenschaften ein antitumorigenes Potenzial besitzt. Um diesen Aspekt genauer zu adressieren, wurde die zytotoxische Wirkung von LA in verschiedenen Dickdarmkrebszellen genauer analysiert und eine mögliche Potenzierung des Zytostatikums 5-Fluorouracil (5-FU) überprüft. In dieser Studie konnten wir zeigen, dass LA in den Tumorzellen vor allem caspaseabhängige, aber auch caspaseunabhängige Zelltodwege einleitet (Abb. 4), wobei diese ohne Beteiligung des Tumorsuppressors und Apoptose-Regulators p53 ablaufen [10]. Interessanterweise ist LA dabei selbst nicht DNA-schädigend. Weiterhin konnte nachgewiesen werden, dass LA die zytotoxischen Effekte des Antimetaboliten 5-FU in diversen Dickdarmkrebszelllinien verstärkt, was LA zu einem möglichen Kandidaten für die Tumortherapie macht.
Ausblick
DNA-Reparaturprozesse spielen eine äußerst wichtige Rolle beim Schutz vor (Darm-)Krebs. Ein genaueres Verständnis der kritischen Reparaturprozesse sowie deren Beeinflussung durch Nahrungsfaktoren erlauben die Identifizierung von Populationen mit einem erhöhten Krebsrisiko, eine verbesserte (Chemo-)Prävention von Krebserkrankungen und die Modulation der Reparaturprozesse im Rahmen einer Krebstherapie. Die Arbeiten werden durch die Deutsche Forschungsgemeinschaft und die Universitätsmedizin Mainz unterstützt.
Literatur
[1] Fahrer, J. & Kaina, B. (2013) Carcinogenesis 34(11), 2435–42
[2] Bastide, N.M. & Pierre, F.H. (2011) Cancer Prev. Res. (Phila) 4(2), 177–84
[3] Wirtz, S. et al. (2010) Carcinogenesis 31(12), 2111–7
[4] Fahrer, J. et al. (2007) Nucleic Acids Res. 35(21), e143
[5] Mangerich, A. & Burkle, A. (2012) Oxid. Med. Cell. Longev. 2012, 321653
[6] Niture, S.K. et al. (2007) Carcinogenesis 28(2), 378–89
[7] Huang, H. et al. (2012) Oncol. Rep. 27(6), 2050–6
[8] Rochette, L. et al. (2013) Mol. Nutr. Food Res. 57(1), 114–25
[9] Göder, A. et al. (2015) Carcinogenesis, in Druck
[10] Dörsam, B. et al. (2014) Arch. Toxicol., in Druck
Foto: © istockphoto.com | Lammeyer
|
L&M 6 / 2015
Diese Artikel wurden veröffentlicht in Ausgabe L&M 6 / 2015.
Das komplette Heft zum kostenlosen Download finden Sie hier:
zum Download
Der Autor:
Weitere Artikel online lesen
News
Mit dem HPLC-Säulenkonfigurator unter www.analytics-shop.com können Sie stets die passende Säule für jedes Trennproblem finden. Dank innovativer Filtermöglichkeiten können Sie in Sekundenschnelle nach gewünschtem Durchmesser, Länge, Porengröße, Säulenbezeichnung u.v.m. selektieren. So erhalten Sie aus über 70.000 verschiedenen HPLC-Säulen das passende Ergebnis für Ihre Anwendung und können zwischen allen gängigen Herstellern wie Agilent, Waters, ThermoScientific, Merck, Sigma-Aldrich, Chiral, Macherey-Nagel u.v.a. wählen. Ergänzend stehen Ihnen die HPLC-Experten von Altmann Analytik beratend zur Seite – testen Sie jetzt den kostenlosen HPLC-Säulenkonfigurator!
© Text und Bild: Altmann Analytik
Aufnahme, Dokumentation und Teilen von Ergebnissen mit ZEISS Stemi 305 und ZEISS Stemi 508
ZEISS stellt zwei neue kompakte Greenough-Stereomikroskope für Ausbildung, Laborroutine und industrielle Inspektion vor: ZEISS Stemi 305 und ZEISS Stemi 508. Anwender sehen ihre Proben farbig, dreidimensional, kontrastreich sowie frei von Verzerrungen oder Farbsäumen.
© Text und Bild: Carl Zeiss Microscopy GmbH
|