

|
Bio&Biotech
 >
Bakterien
>
Bakterien
BakterienArbeiter im MeerBakterien sind Bestandteil des Planktons, ihre durchschnittliche Anzahl liegt bei 109 pro Liter Seewasser, übertrifft also die Zahl aller anderen Organismen um Größenordnungen. Wegen ihrer metabolischen Diversität sind sie essenziell für die Stoffkreisläufe von Kohlenstoff, Stickstoff, Phosphor und Schwefel (auch für Si, Hg, Fe, Mn). 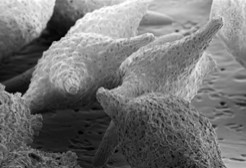
Bis in die 1980er-Jahre wurde die Rolle von Bakterien am Stoffaustausch im Ozean völlig unterschätzt. Dann wurde erkannt, dass Bakterien einen enormen Einfluss auf den Transport von organischem Kohlenstoff ausüben. Der Begriff „mikrobielle Schleife“ beschreibt den Stoffkreislauf, in dem gelöste organische Verbindungen heterotrophen Bakterien als Nahrungsgrundlage dienen. Über verschiedene Organismen wie Flagellaten, Protozoen oder Rotarien werden Bakterien abgeweidet und der Kohlenstoff wieder in die ozeanischen Nahrungsnetze zurückgeführt. Heterotrophe Bakterien setzen nicht nur organisches Material um, sondern stellen auch einen hohen Anteil der organischen Partikel und verbrauchen etwa die Hälfte des durch Fotosynthese fixierten Kohlenstoffs. Der größte Teil des Bakterioplanktons lebt frei im Wasser, bis zu 20% sind an Algen oder an Partikel (Marine Snow) gebunden. Erstaunlich ist die große Artenvielfalt des Bakterienplanktons, sind die Ozeane im Prinzip doch nur riesige, gleichförmige Wasserkörper, arm an Nährstoffen und ohne ökologische Nischen, wie sie an Land anzutreffen sind und dort für Biodiversität sorgen. Zeitliche ökologische Nischen In einer 2012 erschienen Arbeit beschreibt eine Gruppe von 26 Wissenschaftlern vom MPI Bremen, dem Alfred-Wegener-Institut und der Uni Greifswald, wie sich das Bakterioplankton vor, während und nach einer Algenblüte ändert [1]. Die Daten wurden 2009 vor Helgoland gewonnen und die Bakterienpopulationen mithilfe der Fluoreszenz-in-situ-Hybridisierung bestimmt. Vor der Algenblüte dominieren Alfaproteobakterien. Sobald sich aber die Algen massenhaft vermehren, steigt der Gehalt von Flavobakterien (Ulvibacter spp.) innerhalb von wenigen Tagen um das Fünffache an. Ihnen folgen Formosa- und Polari-Bakterien, nachdem die Algenpopulation durch Zooplankton bereits dezimiert wurde und die Ulvibacter-Population abrupt abgebrochen war. Es herrscht also ein stetes „Kommen und Gehen“. Einige Wochen nach der Algenblüte wird wieder der alte Status erreicht. Die Analyse des vollständigen Metagenoms und des Metaproteoms liefert die Erklärung für die zunächst rätselhafte Bakterienfluktuation. Mit so genannten TonB- abhängigen Transporterproteinen werden von Ulvibacter aus dem Algensubstrat zunächst größere Moleküle in das Innere der Zelle befördert. Auch Enzyme für den Abbau komplexer Kohlenhydrate wie Laminarin sind zu diesem Zeitpunkt aktiv. Die späteren Bakterien- populationen müssen sich mit den Resten des „Algenabfalls“, kleineren Peptiden und Zuckern begnügen – nach dem Motto „Wer zuerst kommt, mahlt zuerst“. Das Algensubstrat stellt also offensichtlich ökologische Nischen zur Verfügung, in denen spezialisierte Populationen zeitabhängig ihren Nahrungsbedarf zum Überleben decken können. Bakterien im Stickstoffkreislauf Der maritime Stickstoffkreislauf wird hauptsächlich durch mikrobielle Prozesse gesteuert. Wichtigste Quelle ist die biologische Fixierung von N2 durch Bakterien. Nach mehrfachen Umwandlungsprozessen liegt er in Wasser als Ammonium (NH4+), Nitrit (NO2-), Nitrat (NO3-) und organischer Stickstoff (Norg) vor. Bis zum Beginn der 2000er-Jahre herrschte Konsens über den Prozess der Denitrifikation, mit dem der bakterielle Abbau stickstoffhaltiger Nährstoffe zu N2 im Ozean bezeichnet wird. Die Denitrifikation verläuft je nach Tiefenschicht aerob oder anaerob in Sedimenten unterhalb der sauerstoffhaltigen Zonen und in sauerstoffarmen (cO2 < 10µM/L) tieferen Gewässern (> 300m). Die O2-Minimumzonen (OMZ) machen global nur etwa 0,1% des Ozeanvolumens aus, dennoch sollen hier 30–50% des Gesamtstickstoffs dem System entzogen werden. Die Prozesse laufen dort aber nicht über die Denitrifikation, sondern über einen neuen mikrobiellen Stoffwechselweg, der Anammox-Reaktion (Anaerobe Ammonium Oxidation) von NH4+ mit NO2- zu N2 durch Anammox-Bakterien (Abb.1). Die Gesamtreaktion ist im Prinzip eine Symproportionierung; als Intermediate treten dabei die toxischen Moleküle Hydrazin und Hydroxylamin auf.

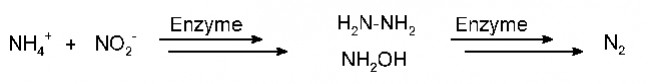
Der Anammox-Prozess Seit ihrer taxomomischen Identifizierung wurden vier Gattungen von Anommox-Bakterien gefunden: Candidatus Brocadia, Candidatus Kuenenia, Candidatus Anammoxoglobus und Candidatus Scalindua. Anammox-Bakterien wurden erstmals in Kläranlagen in Holland, später von Wissenschaftlern des MPI für Marine Biologie Bremen um M. S. Jetten auch erstmals im Schwarzen Meer entdeckt. Inzwischen weiß man, dass sie weltweit in OMZ-Regionen vorkommen. Bakterien, die zur Anammox-Reaktion befähigt sind, enthalten eine spezielle Organelle, das Anammoxoson, in dem die Reaktion stattfindet. Schlüsselenzym ist eine Hydroxylamin-Oxidoreduktase, mit deren Hilfe das intermediär entstandene Hydrazin und Hydroxylamin zu N2 oxidiert werden. Die Bakterienmembran ist einzigartig aufgebaut, sie besteht aus Ladderan-Lipiden, die drei bzw. fünf annelierte Cyclobutane enthalten (Abb. 2) und als Ester oder Ether an die Glycerineinheit gebunden sind. Im Vergleich zu den üblichen Membranen zeigen die Lipide eine ungewöhnlich hohe Dichte bis zu 1,6kg/dm gegenüber 1,0kg/dm. Damit können die toxischen Moleküle das Organell nicht verlassen, der Konzentrationsgradient kann aufrechterhalten werden.
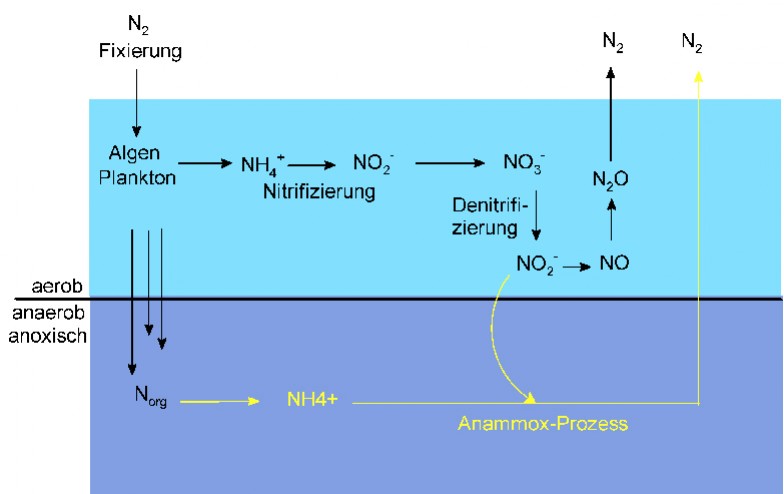
Abb. 1 Stickstoffkreislauf im Meer, schematisch. Die Konzentration von O2 nimmt mit zunehmender Wassertiefe ab. Der Anammox-Prozess, der völlig ohne Sauerstoff verläuft, ist an die Denitrifizierung und die Bildung von NH4+ aus organischem Stickstoff gekoppelt
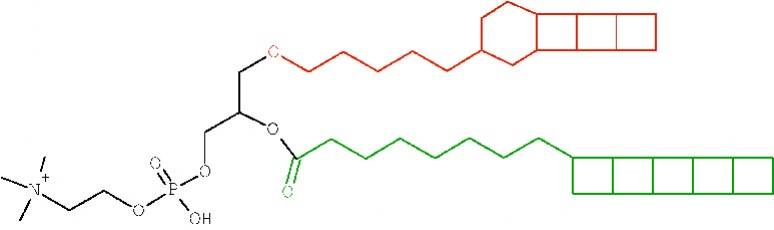
Abb.?2?Prinzipielle Struktur eines Ladderan-Lipidmoleküls mit der polaren Phosphatidylcholingruppe, dem Glycerin-Rückgrat, dem Ladderan-Alkohol (rot) und der Ladderan-Fettsäure (Pentacycloanammoxinsäure, PCAS)
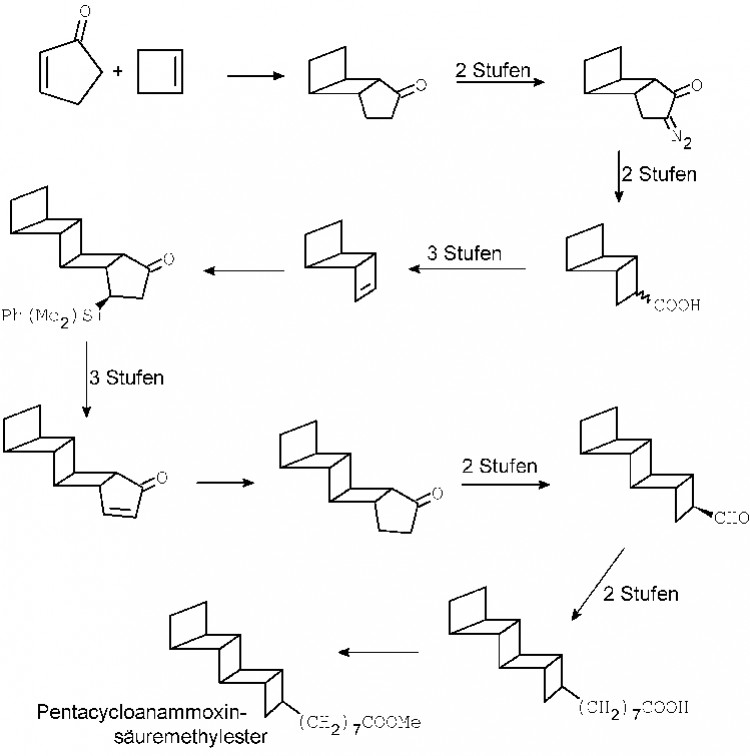
Abb. 3 Schematische Darstellung des von E. J. Corey et al. gewählten Synthesewegs zum Pentacycloanammoxinsäuremethylester
Die ungewöhnliche Ladderanstruktur und die Schwierigkeiten, C.B. anammoxidans zu kultivieren und damit genügend Substanz zu erhalten, sind natürlich Herausforderungen, die Synthese dieser Verbindungen anzugehen. Über den Ladderanalkohol sind unseres Wissens keine Daten bekannt, die erste Synthese der razemischen Pentacycloanammoxinsäure (PCAS) wurde von E. J. Corey beschrieben [2]. Die Synthese verläuft über 17 Stufen mit einer Gesamtausbeute von 0,25%. Immerhin konnte damit die Struktur, aber nicht die absolute Konfiguration bewiesen werden. 2006 erschien dann die asymmetrische Synthese (Abb.?3) der PCAS [3]. Wieder waren 17 Reaktionsstufen bis zum Zielmolekül erforderlich, die Ausbeute betrug lediglich 1,9%. PCAS ist wegen der hohen Spannungsenergie der Cyclobutanringe ziemlich instabil. Der Anammox-Prozess ist eine echte Alternative zur klassischen Entfernung von Stickstoffverbindungen in Kläranlagen. Die Kosten reduzieren sich auf etwa 10% und der Ausstoß von CO2 reduziert sich um über 85% gegenüber konventionellen Anlagen. Die weltweit erste Anlage ging schon 2000 in Hattingen in Betrieb, 2004 folgte Gelsenkirchen, dann Rotterdamm 2006.
Literatur Foto: ©Panthermedia.net|stephan kerkhofs |
L&M 4 / 2013
Das komplette Heft zum kostenlosen Download finden Sie hier: zum Download Der Autor:Weitere Artikel online lesen


NewsSchnell und einfach die passende Trennsäule finden
© Text und Bild: Altmann Analytik ZEISS stellt neue Stereomikroskope vor
ZEISS stellt zwei neue kompakte Greenough-Stereomikroskope für Ausbildung, Laborroutine und industrielle Inspektion vor: ZEISS Stemi 305 und ZEISS Stemi 508. Anwender sehen ihre Proben farbig, dreidimensional, kontrastreich sowie frei von Verzerrungen oder Farbsäumen. © Text und Bild: Carl Zeiss Microscopy GmbH |






