


|
Umweltfaktoren in besonders empfindlichen Phasen der Entwicklung
Umweltfaktoren in besonders empfindlichen Phasen der EntwicklungDas Risiko aus der UmweltOrganismen sind in jeder Lebensphase, von der Empfängnis bis zu ihrem Tod, einer Vielzahl von Umweltfaktoren ausgesetzt, die für sie teilweise mit hohem Gesundheitsrisiko verknüpft sind. In aktuellen Forschungsarbeiten soll aufgeklärt werden, welche Bedeutung besonders empfindliche Phasen der Organentwicklung („Developmental Windows of Disease“) besitzen. In diesen Zeitfenstern besteht die Möglichkeit, genetisch determinierte Entwicklungsvorgänge derart zu verändern, dass durch die resultierenden Umprogrammierungen sich das Risiko etwa für metabolische Erkrankungen, für eine eingeschränkte Reproduktionsfähigkeit oder gar eine Tumorgenese im späteren Leben bzw. in der Nachkommenschaft verändert. 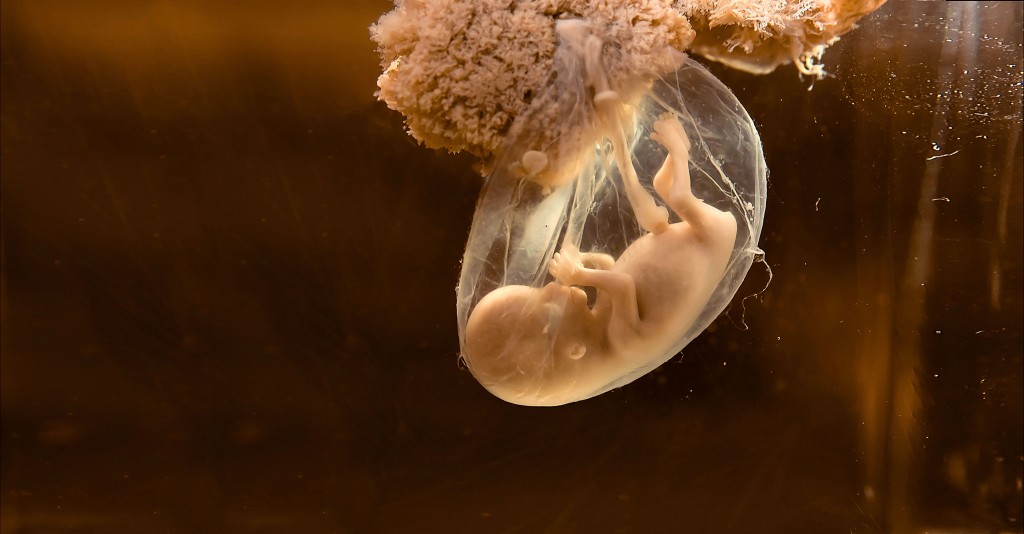
Die Individualentwicklung eines Säugetieres vom frühen Fötus bis ins Erwachsenenalter ist genetisch determiniert. Der Ablauf dieses Programms unterliegt aber zusätzlich dem Einfluss zahlreicher Umwelteinflüsse, wobei speziell Toxine, Stress- bzw. Verhaltensfaktoren sowie die Ernährung hervorgehoben werden sollen (Abb.1). Die Nahrung nimmt in dreierlei Hinsicht Einfluss, wobei Hunger und Überernährung, besonders wenn diese zu Übergewicht führt, eher generellere Einflüsse darstellen, während Lebensmittelinhaltsstoffe in der Regel spezifisch auf individuelle molekulare Strukturen wie z.B. Rezeptoren oder Enzyme einwirken. Der Einfluss von Umweltfaktoren auf ablaufende entwicklungsbiologische Programme ist so stark, dass er die Gesundheit des betroffenen Individuums lebenslang beeinflussen kann. Über epigenetische Mechanismen wird ein Teil der Umwelteinflüsse vererbbar, sodass auch die Gesundheit weiterer Folgegenerationen beeinflusst werden kann, dem so genannten „Developmental Programming of Disease“.

Abb.1 Umweltfaktoren und Gesundheit: Umweltfaktoren wie Stress, Toxine, Veränderungen im Verhalten und bei der Ernährung sind in der Lage, über die Modulation entwicklungsbiologischer Abläufe die Gesundheit im späteren Leben oder die Gesundheit von Nachfolgegenerationen zu beeinflussen. Dabei scheint insbesondere der Modulation von epigenetischen Mechanismen eine wichtige Rolle zuzukommen. Bezüglich des Einflusses der Ernährung muss einerseits zwischen Effekten, ausgelöst durch die Verfügbarkeit und die (kalorische) Qualität von Nahrungsmitteln (Hunger/Überfluss), und Effekten durch Lebensmittelinhaltsstoffe unterschieden werden, die wie Pharmaka auf sehr spezifische molekulare Zielstrukturen einwirken wie z.?B. Rezeptoren oder Enzyme.
Umwelteinflüsse haben Auswirkung auf mehrere Generationen Mechanistisch betrachtet scheinen Umwelteinflüsse vor allem über epigenetische Mechanismen entwicklungsbiologische Prozesse umzuprogrammieren (zur Übersicht siehe Gerhäuser [1]). Dies kann Folgen für die direkt betroffene Generation, aber auch für die darauf folgenden haben, obwohl diese dem Umwelteinfluss, etwa einer Chemikalie, nicht mehr direkt ausgesetzt waren. In einem schwangeren/trächtigen Organismus können im ungünstigsten Fall gleichzeitig die Mutter, die Nachkommen und auch deren Keimzellen, also die F0-, F1- und F2-Generation, direkt betroffen sein. Prägt sich der entstehende Effekt neben der Mutter in der F1- und/oder der F2-Generation aus, spricht man von einem Multigenerationseffekt. Persistiert dieser Effekt in der F3- oder späteren Generation, also in Generationen ohne direkten Chemikalieneinfluss, spricht man von einem Transgenerationseffekt. Beide, Multi- und Transgenerationseffekte, konnten z.B. in tierexperimentellen Studien unzweifelhaft für sogenannte. „Endocrine Disrupting Chemicals (EDC)“, bezogen auf eine eingeschränkte männliche Reproduktionsfähigkeit, gezeigt werden. Dabei wurde das Epigenom der Keimbahnzellen umprogrammiert und die dadurch veränderten entwicklungsbiologischen Prozesse als epigenetische Veränderungen über die männliche Keimbahn an männliche Nachkommen weitervererbt [2,3]. Daraus ergibt sich die Frage, ob derartige Mechanismen auch für den Menschen relevant sind. An den Folgen des holländischen Hungerwinters von 1944/1945 konnte dies belegt werden. Wissenschaftler der Universität Leiden analysierten Schicksale der damaligen Schwangeren und ihrer Kinder. Anhand der Ergebnisse wissen wir, dass die Töchter der von Hunger betroffenen Mütter ein klar erhöhtes Risiko besitzen z.B. an einer Stoffwechselerkrankung (darunter Fettsucht, erniedrigter Glukosetoleranz, Bluthochdruck), an neuronalen Störungen oder an hormonabhängigen Tumorerkranken wie z.B. der Brustdrüse zu erkranken. Dieser Effekt bleibt auch in der Enkelgeneration bestehen. Diese Beispiele belegen, dass Umwelteinflüsse (hier Verfügbarkeit von Nahrung), die auf entwicklungsbiologische Abläufe einwirken, das Risiko für Erkrankungen später im Leben und in nachfolgenden Generationen beeinflussen können. Inzwischen gibt es zusätzlich aus epidemiologischen Erhebungen und tierexperimentellen Studien erste Hinweise darauf, dass Inhaltsstoffe aus Nahrungsmitteln über die Modulation entwicklungsbiologischer Abläufe auch zur Prävention von Erkrankungen führen könnten [4,5]. Relevante biologische Zeitfenster In Anbetracht der Kenntnis, dass Umweltfaktoren den Ablauf entwicklungsbiologischer Vorgänge umprogrammieren können, stellt sich die Frage des Timings. Reagiert dabei der Organismus zu jeder Phase seiner Entwicklung von der Empfängnis bis ins Erwachsenenalter gleich empfindlich oder gibt es besonders empfindliche und fragile Zeitfenster? Bei der Aufarbeitung der Folgen des oben erwähnten Hungerwinters zeigte sich, dass die beobachteten Effekte, ähnlich wie bei teratogenen Effekten, z.B. durch Thalidomid verursacht, mit definierten Phasen der Schwangerschaft korrelieren, in denen die Mutter hungerte. Das erhöhte Risiko des Kindes für Fettsucht korrelierte mit einer Hungerperiode der Mutter im ersten Trimester der Schwangerschaft, das Risiko für eine erniedrigte Glukosetoleranz war mit Hungern im mittleren Drittel assoziiert und das Risiko für Bluthochdruck entstand, wenn die Mutter im letzten Trimester hungerte [4,5]. Insgesamt kann festgehalten werden: a) Es gibt besonders fragile Zeitfenster für die Programmierung von gesundheitlichen Konsequenzen durch Umweltfaktoren in frühen Phasen der Entwicklung und b) diese Zeitfenster der Fragilität scheinen organ- oder prozessspezifisch zu sein. Der größte Risikofaktor für die Entstehung eines malignen Tumors der Brustdrüse scheint die lebenslänglich akkumulierende Menge an körpereigenen Östrogenen und aufgenommenen östrogenartig wirkenden Substanzen zu sein. Die Belastung mit diesen Hormonen scheint allerdings nicht zu jeder Phase im Leben einer Frau ein ähnlich hohes Risiko darzustellen. Als kritische Phasen werden beim Menschen bisher für die frühe fötale Entwicklung, die pubertäre Entwicklung der Brustdrüse, die Involution der Brustdrüse infolge des Abstillens bzw. des Eintritts der Menopause diskutiert (Abb.2 und [6]). Aus tierexperimentellen Studien ist aber bekannt, dass eine weitere kritische Phase die neonatale Entwicklung der Brustdrüse umfasst.
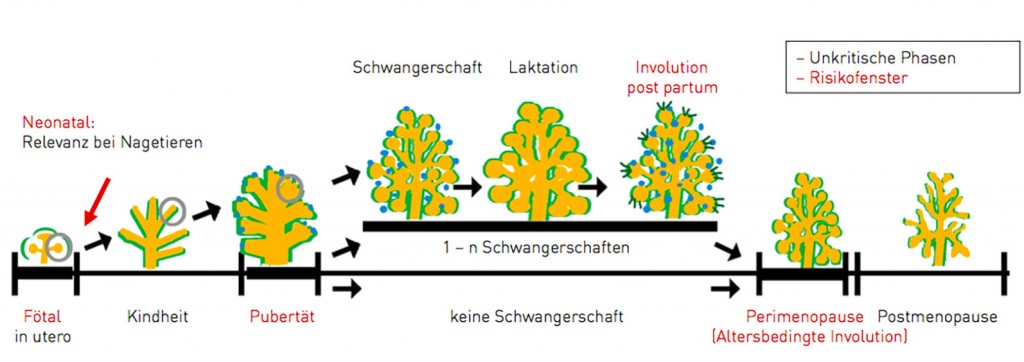
Abb.2 Hypothese zu Risikofenstern für die Entwicklung von Brustkrebs. Schematische Darstellung des Lebenszyklus der Entwicklung der Milchdrüse der Frau. Der Schwangerschafts-, Laktations- und Involutionszyklus der Brust unterscheidet Mütter von Frauen, die nie schwanger waren. Die vier Zeitfenster der besonderen Relevanz hinsichtlich des Brustkrebsrisikos sind rot beschriftet und umfassen Teile der fötalen und pubertären Entwicklung, sowie Involutionsprozesse ausgelöst durch Abstillen oder sinkender Hormonproduktion durch die Ovarien. Modifiziert nach [6].
Xenoöstrogene aus der Umwelt Wegen dieser Einflüsse werden sogenannte „Umweltöstrogene“ besonders intensiv untersucht. Diese Xenobiotika mit östrogenartiger Wirkung umfassen sowohl Industriechemikalien als auch pflanzliche und Pilzsubstanzen (Myco- u. Phytoöstrogene). Aus pflanzlichen Quellen, die besonders reich an Phytoöstrogenen sind, werden deshalb Auszüge (Extrakte) erstellt und kommerziell als Ersatzstoffe für die Behandlung von Wechseljahresbeschwerden zum Verkauf angeboten. Hierzu gehören u.?a. Isoflavone aus Soja, Rotklee und Kudzu sowie Derivate des Naringenins aus dem Hopfen. Derartige Extrakte bzw. deren Inhaltsstoffe werden in unserer Arbeitsgruppe hinsichtlich ihrer Wirksamkeit (z.B. Verhinderung von Osteoporose, Unterdrückung vasomotorischer Störungen) und Sicherheit (Ausschluss der Risiken für die Brustkrebsentwicklung bzw. Hyperplasien des Endometriums) in präklinischen experimentellen Modellen untersucht. Wie oben ausgeführt, beschäftigen wir uns seit vielen Jahren mit der hormonellen Wirksamkeit von Inhaltsstoffen aus der Nahrung und aus Heilpflanzenpräparaten, u.a. mit Isoflavonen aus Leguminosen. Doch wie kommt man, ausgehend von Untersuchungen zur molekularen und zellulären Wirkung von Sojapräparaten und deren Inhaltsstoffen, dazu, sich mit Hochrisikofenstern der Brustdrüsenentwicklung zu beschäftigen? Die Idee zum Einsatz von Sojapräparaten für die Behandlung von hormonbedingten Syndromen wie Wechseljahresbeschwerden hat ihren Ursprung in Beobachtungen von Prof. Adlercreutz [7]. Er konnte beim Vergleich europäischer und asiatischer Probandenkollektiven zeigen, dass sich der Spiegel von Isoflavonen (den wirksamen Inhaltsstoffen in Soja) im Urin umgekehrt proportional zum Brustkrebsrisiko in diesen Gruppen verhält. Der kausale Zusammenhang zwischen der Menge der aufgenommenen Sojaprodukte und der Erniedrigung des Brustkrebsrisikos konnte aber erst im Zeitraum zwischen 2009 und 2014 anhand klinischer Daten erhärtet werden [8]. Eine lebenslange Aufnahme von Isoflavonen aus Soja über die Nahrung scheint also mit einem niedrigeren Risiko verbunden zu sein, an Brustkrebs zu erkranken. Klinische Studien zur Anwendung von mit Sojaisoflavon angereicherten Präparaten bei Frauen in den Wechseljahren haben zwar bislang keine Hinweise auf Nebenwirkungen wie z.B. der Veränderung des Brustdrüsengewebes erbracht, allerdings ergaben verschiedene tierexperimentelle Untersuchungen, dass Sojaisoflavone durchaus in der Lage sind, das Wachstum von humanen Mammakarzinomzellen, die auf immundefiziente Nacktmäuse übertragen wurden, zu fördern [9]. Übersetzt man obige Erkenntnisse in eine pharmakologische Sprachweise, so entspricht eine lebenslängliche Aufnahme von Isoflavonen aus Soja einer chronischen, niedrigdosierten Exposition, die sich in einer fortgeschrittenen Lebensphase begünstigend auf das Brustkrebsrisiko auswirken könnte. Die Aufnahme von Isoflavonen als angereichertes Nahrungsergänzungsmittel zum Zeitpunkt des Klimakteriums hingegen entspräche dann einer relativ hochdosierten Behandlung zu einem späten Lebensabschnitt. Aus diesen Beobachtungen und der Hypothese, dass es auch für die Entwicklung von Tumoren der Brustdrüse besonders sensible und damit kritische Zeitfenster gibt, leiteten sich die wissenschaftlichen Fragestellungen ab, die in den letzten Jahren im Zentrum der Forschungsaktivitäten unserer Arbeitsgruppe standen: a) Wie wirkt sich im experimentellen Modell der Ratte eine lebenslang verabreichte, sojareiche Nahrung auf die Wirkung des weiblichen Sexualhormons Östradiol als Triebfeder der Karzinogenese hormonabhängiger Organe aus und b) wie wirkt sich eine eventuelle Modulation der Östradioleffekte auf die Genese eines östradiolgetriebenen, experimentellen Modells von Mammakarzinomen aus? Isoflavone modulieren Wirksamkeit von Östradiol Da die Wirkung von Östradiol eine entscheidende Bedeutung für die Karzinogenese hormonabhängiger Organe besitzt, führten wir einen ersten generationenübergreifenden Fütterungsversuch mit Tierfutter durch, das definierte Mengen an Isoflavonen aus Soja enthielt. Kontrolltiere erhielten ein phytoöstrogenfreies Kontrollfutter. Die Verfütterung dieser Diäten wurde in der Elterngeneration vor der Verpaarung begonnen und in der Nachkommengeneration über alle entwicklungsbiologisch kritischen Zeitfenster (fötale, neonatale und pubertäre Phase) aufrechterhalten. Mit einem sogenannten Uterotrophietest prüften wir am postnatalen Lebenstag 97, ob und wie sich die Isoflavone aus dem Futter auf die Ansprechbarkeit der Zielorgane Gebärmutter und Brustdrüse auf Östradiol auswirken. Die Schlüsselergebnisse bestanden darin, dass die Empfindlichkeit des Uterusgewebes auf eine Östradiolbehandlung um ein Vielfaches gesteigert wurde [10], während die östradiolabhängige Stimulation der Proliferation des Brustdrüsenepithels durch Isoflavone klar gedämpft war [11]. Letzteres ist von eindeutiger Relevanz für die Bewertung des Einflusses von Sojaisoflavonen auf das Brustkrebsrisiko. Da in den Tieren, die Futter erhielten, welches mit Isoflavonen angereichert war, auch noch die Knochenmasse und -dichte erhöht waren, können wir die Ergebnisse dieser Untersuchungen folgendermaßen zusammenfassen: a) Die kontinuierliche Sojaisoflavoneexposition programmiert die Ansprechbarkeit hormonabhängiger Organe für das weibliche Sexualhormon Östradiol um und b) die Auswirkung der Beeinflussung entwicklungsbiologischer Prozesse durch Umwelteinflüsse ist organselektiv, wie die gegensätzlich ausgeprägten Effekte in den beiden Hauptzielorganen für Östradiol, Gebärmutter und Brustdrüse, zeigen. Modulieren Isoflavone auch die Karzinogenese der Brustdrüse? Die Karzinogenese der Brustdrüse ist zumindest in ca. 70% der Fälle ein östradiolabhängiger Prozess. Das gilt sowohl für die Initiation der Karzinogenese durch genotoxische Östradiolmetaboliten als auch für die Promotion der Karzinogenese des Mammakarzinoms durch Östradiol in einem Rezeptor vermittelten Prozess. Aus diesem Grund ist unser Befund, dass Isoflavone die Östradiolempfindlichkeit des Brustdrüsengewebes modulieren, von hoher Relevanz. Das Ziel unserer aktuellen Arbeiten ist es deshalb zu prüfen, ob durch Isoflavone auch die östrogenabhängige Karzinogenese der Brustdrüse beeinflusst wird. Um dies zu klären, bedarf es eines geeigneten Modells, in dem östradiolabhängig Mammakarzinome entstehen. Für diesen Zweck eignen sich insbesondere weibliche ACI-Ratten. Erhöht man bei diesen Tieren künstlich die endogenen Östradiolspiegel, dann entstehen mit hoher Inzidenz Brustdrüsentumore [12]. Dieses langwierige Tumorexperiment hat Dr. Frank Möller in Kooperation mit Dr. Oliver Zierau in unserer Arbeitsgruppe durchgeführt. Technisch haben sie das Experiment an unseren oben beschriebenen Fütterungsversuch mit isoflavonhaltigem Futter im Vergleich zu isoflavonfreien Placebofutter angelehnt. Zum Zeitpunkt der Pubertät wurde ein Östradiolpellet eingepflanzt und das Experiment bis zum postnatalen Tag 285 durchgeführt, wobei der Verlauf des Prozesses mithilfe sogenannter Wholemount-Präparate verfolgt werden kann (Abb.3).
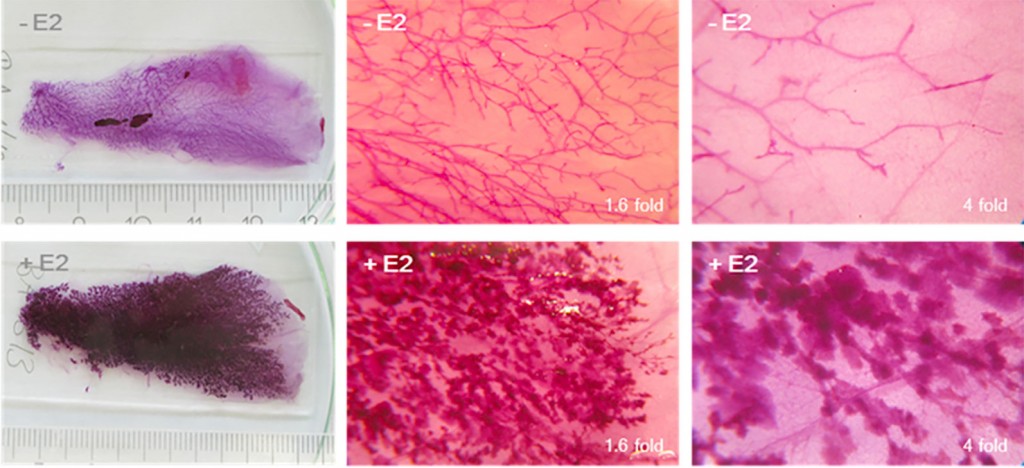
Abb.3 Wholemount-Präparate als Indikatoren für Veränderungen in der Milchdrüse. Untersuchungen mit experimentellen Tumormodellen sind langwierig. Mögliche Veränderungen der Brustdrüse durch die experimentellen Bedingungen können in vielen Fällen relativ frühzeitig in Wholemount-Präparaten des Organs beobachtet werden. Vergleichend dargestellt sind Drüsengänge in Präparaten der Milchdrüse mit und ohne Östradiolstimulation (+E2/-E2). Die Abbildungen wurden von Dr. Frank Möller, Molekulare Zellphysiologie und Endokrinologie, TU-Dresden zur Verfügung gestellt.
Das vielversprechende Experiment ist noch nicht fertig ausgewertet und daher sind noch keine finalen Daten im Detail verfügbar. Als extrem wichtiges Ergebnis können wir aber jetzt schon eindeutig festhalten: Eine Exposition mit Sojaisoflavonen über alle empfindlichen Phasen der Brustdrüsenentwicklung hinweg, moduliert die östradiolabhängige Karzinogenese.
Literatur Bild: © istockphoto.com|Leptospira |
L&M 3 / 2015
Das komplette Heft zum kostenlosen Download finden Sie hier: zum Download Der Autor:Weitere Artikel online lesen


NewsSchnell und einfach die passende Trennsäule finden
© Text und Bild: Altmann Analytik ZEISS stellt neue Stereomikroskope vor
ZEISS stellt zwei neue kompakte Greenough-Stereomikroskope für Ausbildung, Laborroutine und industrielle Inspektion vor: ZEISS Stemi 305 und ZEISS Stemi 508. Anwender sehen ihre Proben farbig, dreidimensional, kontrastreich sowie frei von Verzerrungen oder Farbsäumen. © Text und Bild: Carl Zeiss Microscopy GmbH |





