


|
Zelluläre Transportproteine und Arzneistofftransport
Zelluläre Transportproteine und ArzneistofftransportDie Stärke von Arzneimitteln hängt oft von der Konzentration des Wirkstoffes an seinem Zielort ab. Meistens werden Arzneimittel aber diesem Zielort von fern zugeführt. Der Wirkstoff muss zunächst in Lösung gehen und lokale Barrieren wie die Darmwand durchqueren, um in die Blutbahn und dann an seinen Zielort gelangen zu können. Dabei galt lange Zeit das Dogma, dass die Diffusion des Wirkstoffs die treibende Kraft der Aufnahme in den Körper oder in eine Zelle ist. Er sollte möglichst keine Ladung tragen und lipophil sein, um leicht durch Membranen diffundieren zu können. 
Heute wissen wir, dass der größere Teil von Wirkstoffen, aber auch viele Nahrungsbestandteile, in ionisierter Form über Transportproteine durch Membranen transportiert werden. Transportproteine sind Proteine, die in einer Zellmembran lokalisiert sind und die Permeation einer Substanz durch die Membran passiv oder aktiv vermitteln. Im Gegensatz zur freien Diffusion ist die transportervermittelte Permeation sättigbar und hemmbar und kann sowohl passiv entlang eines Konzentrationsgefälles als auch aktiv gegen einen Konzentrationsgradienten unter Energieverbrauch erfolgen. Die Energie kann entweder direkt vom Transportprotein verbraucht werden (primär aktive Transporter) oder durch indirekte Kopplung eines Transporters an ein primär aktives Protein (z.B. eine ATPase). Wir unterscheiden Uniportsysteme, die Substratmoleküle in eine Richtung transportieren, Symport- oder Kotransportsysteme, die gleichzeitig zwei oder drei verschiedene Substanzen in die gleiche Richtung transportieren oder Antiportsysteme, die zwei oder drei Substanzen in gegensätzliche Richtung transportieren (Abb.1). Daneben gibt es Ionenpumpen (z.B. Na+/K+-ATPasen), Ionenkanäle und Wasser transportierende Aquaporine. Die Aufklärung des Genoms führte zu einer international verbindlichen Klassifizierung der Transportproteine und Ionenkanäle durch die International Union für Biochemie und Molekularbiologie (IUBMB) in der „Transporter Classification database” (TCDB), die mehr als 600 Familien von Transportproteinen einschließt. Die humanen Transportproteine umfassen die Superfamilien der Annexine, ATPasen, Kalziumkanäle, Kaliumkanäle, Natriumkanäle, ATP-bindenden Transportproteine und der Human Solute Carriers (SLCs). Eine Liste aller Gene, die für diese Proteine kodieren, ist auf der Internetseite des HUGO Gene Nomenclature Committee (HGNC) zu finden (www.genena

Abb.1 A) unidirektionaler Transport, Cotransport, Antiport-System. B) sekundär aktiver Transport: Kopplung eines Na+-Aminosäure- oder Gluosetransporters in einer apikalen Membran an eine Na/K-ATPase in der basolateralen Membran. Die nötige Energie erfolgt durch Hydrolyse von ATP durch die ATPase (aus Biopharmazie, VCH-Wiley; mit Genehmigung der Herausgeber). Arzneistoff-Interaktionen auf Transporterebene Eine Reihe von Arzneistoffen vermag Transportproteine zu hemmen und in der Folge die Pharmakokinetik anderer Transportsubstrate zu verändern, was sogar schon dazu führte, dass Medikamente wieder vom Markt genommen wurden. Ein bekanntes Beispiel ist Cerivastatin, ein HMG-CoA Reduktasehemmer, dessen Einnahme in mehreren Fällen zu tödlicher Myotoxizität führte. Eine Analyse ergab, dass einige Patienten gleichzeitig antihyperlipidämisches Gemfibrozil eingenommen hatten, wodurch die Plasmakonzentration von Cerivastatin um das bis zu 4.4-Fache angestiegen war [1]. Ursache war eine Hemmung des CYP2C8-vermittelten Metabolismus und eine Hemmung der OATP2 (organische Anionen transportierendes Protein2, OATP2/OATP1B1:SLC21A6)) vermittelten hepatischen Aufnahme von Cerivastatin durch Gemfibrozilglucuronid [2]. Um solchen Fällen vorzubeugen, erließ die amerikanische FDA 2012 die Richtlinie „Drug Interaction Studies, Study Design, Data Analysis, Implications for Dosing, and Labeling Recommendations“. Im gleichen Jahr wurde die Richtlinie der EMA „Guideline on the Investigation of Drug Interactions“, die speziell transportervermittelte Wirkstoff/Wirkstoff-Interaktionen diskutiert, finalisiert.
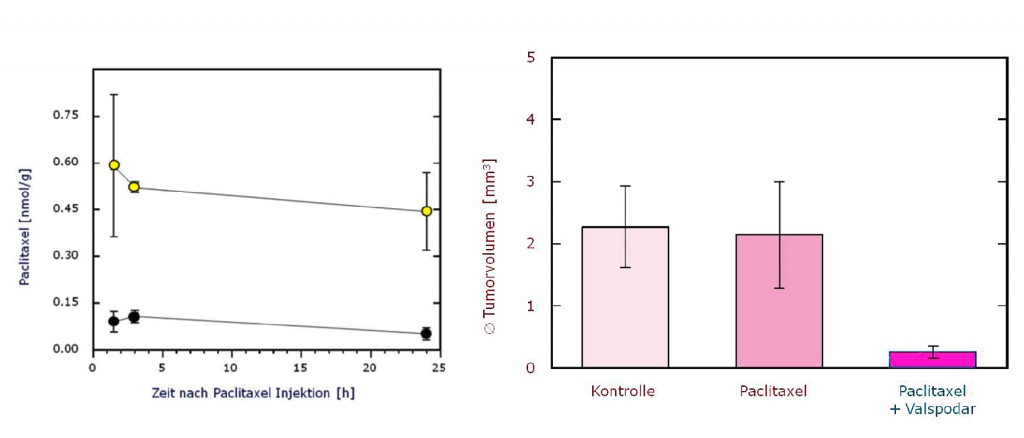
Abb.2 A) Akkumulation des P-Glykoproteinsubstrats Paclitaxel im Gehirn ohne und nach peroraler Verabreichung des P-Glykoproteininhibitors Valspodar (PSC-833). B) Glioblastomgröße 35 Tage nach Tumorimplantation in Kontrolltieren, nach Behandlung mit Paclitaxel allein und nach peroraler Gabe des p-Glykoproteininhibitors Valspodar kurz vor der i.v. Verbreichung von Paclitaxel. ATP-bindende Proteine Gemeinsames Kennzeichen der Superfamilie der ABC (ATP binding cassette)- Transporter ist ihre Fähigkeit, ATP zu binden, zu hydrolysieren und damit die Energie für die Translokation von Substraten zu gewinnen. ABC-Transporter agieren unidirektional entweder als Import- oder Exportproteine und sind in Lebewesen aller Entwicklungsstufen zu finden. Während ABC-Exportproteine in Prokaryonten und Eukaryonten exprimiert werden, wurden ABC-Importproteine bislang ausschließlich in Prokaryonten nachgewiesen. ABC-Transporter stellen mit ca. 80 Systemen die umfangreichste Proteinfamilie in E. coli und repräsentieren ca. 5% des bakteriellen Genoms, während bislang 48 ABC-Transporter in sieben Familien im Menschen beschrieben sind. Die sieben dafür kodierenden Genfamilien werden in moderner Nomenklatur von ABCA bis ABCG bezeichnet. Diese Transporter sind am Lipid- und Cholesterintransport, an der Antigenpräsentation, an der mitochondrialen Eisenhomöostase, der ATP-abhängigen Regulation von Ionenkanälen und der Resistenz gegen Chemotherapeutika beteiligt. Besondere Bedeutung besitzen die ABC-Exportpumpen P-Glykoprotein (MDR1, P-GP; ABCB1) und Breast Cancer Resistance Protein (ABCG2) an der Blut-Hirn-Schranke, wo sie eine wesentliche Komponente der Barriere zum Schutz des zentralen Nervensystems darstellen. Leider werden dadurch viele ZNS-Erkrankungen schwer behandelbar, weil eben auch viele potenziell hochwirksame Wirkstoffe nicht ins Gehirn eindringen können. Besonders offensichtlich wird dies bei der Behandlung von Gehirntumoren oder Metastasen, bei denen die Anwendung von Zytostatika meist wirkungslos bleibt. So kann beispielsweise das Mitosegift Paclitaxel (Taxol), das erfolgreich bei Mammakarzinom oder Ovarialkarzinomen angewandt wird, nicht zur Behandlung von Glioblastomen eingesetzt werden, weil es als p-Glykoproteinsubstrat die Blut-Hirn-Schranke nicht durchquert. Tierversuche in Ratten mit einem implantierten humanen Glioblastom haben eindrucksvoll gezeigt, dass die Hemmung von P-Glykoprotein in der Blut-Hirn-Schranke durch einen gleichzeitig verabreichten Blocker zu einer drastischen Erhöhung der Paclitaxelkonzentration im Gehirn (Abb. 2) und in der Folge zu einer deutlichen Tumorreduktion führen kann [4]. Transportproteine und Krankheiten Transportproteine können auch unmittelbar an Krankheiten beteiligt und Defekte an Transportern Krankheitsursache sein: Der Glucosetransporter SLC2A2 (GLUT-2) wird vor allem in Leber, Pankreas und Dünndarm und Niere exprimiert und ermöglicht insulinunabhängig den Transport von Glucose. Mutationen im SLC2A2-Gen verursachen das seltene Fanconi-Bickel-Syndrom, eine Zuckerspeicherkrankheit. Die Glutamattransporter SLC1A2 und SLC1A3 sollen in der Pathogenese von amyotropher Lateralsklerose, Alzheimer Krankheit, Autismus und Schizophrenie involviert sein. Aus der Familie der SLC13-Transporter (Na+-gekoppelte Di- und Tricarboxylat/Sulfattransporter) sollen SLC13A2 (NaDC1) und SLC13A3 (NaDC3) an der Entstehung von Nierensteinen und der Pathogenese der Stoffwechselkrankheiten Glutarazidurie Typ1 und Canavan Disease beteiligt sein [5]. Dem Anionentransporter SLC26A4 wird eine Rolle bei Taubheit zugeschrieben, SLC31A1 und SLC31A2 bei der Kupferspeicherkrankeit Wilsons Disease. Zu den am besten untersuchten genetisch bedingten Erkrankungen zählt die zystische Fribrose oder Mukoviszidose, als deren Ursache Mutationen am langen Arm des Chromosoms 7 gelten. Die Mukoviszidose ist die zweithäufigste angeborene Stoffwechselerkrankung hellhäutiger Menschen, wobei die Erkrankungsquote bei etwa 1:2.000 Neugeborenen liegt. Das betroffene Gen codiert für CFTR (Cystic Fibrosis???Transmembrane Conductance Regulator (ABCC7)), einen Chloridkanal, der zu den ABC-Transportern zählt. Bei einer Störung dieses Kanals ist der Wassergehalt von Sekreten der Bronchien, des Pankreas, der Leber und des Dünndarms zu niedrig, und sie werden zähflüssig, wodurch es zu unterschiedlichsten Funktionsstörungen der betroffenen Organe kommt. Bisher sind über 1.000 verschiedene Mutationen des CFTR-Gens bekannt. Die häufigste Mutation dieses Gens, ? F508, betrifft das Fehlen von Phenylalanin an der Position 508 im Protein, sie tritt bei sieben von zehn Menschen mit Mukoviszidose auf. Eine weitere, allerdings seltene Erbkrankheit, die auf einen ABC-Transporterdefekt zurückzuführen ist, ist die Tangier-Krankheit, eine Erkrankung des Fettstoffwechsels, der ein Defekt des ABCA1-Gens zu Grunde liegt, das für einen Cholesterintransporter kodiert. Dadurch kommt es zur verringerten Bildung von High Density Lipoproteinen und zu einer verstärkten Cholesterinspeicherung. Bekannter ist das autosomal rezessive Dubin-Johnson-Syndrom, das durch eine Mutation des Gens für das multidrug resistance related protein 2 (MRP2, ABCC2) bedingt wird. Durch das Protein wird glucuronidiertes (konjugiertes) Bilirubin ATP-abhängig von der Leber in die Galle sezerniert. Bei Funktionslosigkeit des Proteins kann Bilirubin nicht in die Gallenkapillaren transportiert werden, was zu einer Anreicherung in der Leber und zum Rückstau konjugierten Bilirubins in das Blut führt. Glücklicherweise hat die Erkrankung eine gute Prognose und eine Behandlung ist normalweise nicht notwendig. Mutationen im ABCB1-Gen, das den bekanntesten ABC-Transporter – P-Glykoprotein (ABCB1) – kodiert, sollen zu erhöhter Anfälligkeit für einen Subtyp chronisch-entzündlicher Darmerkrankungen (Inflammatory bowel disease 13) führen. Möglicherweise ist die Exportpumpe auch zusammen mit ABCG2 (breast cancer resistant protein, BCRP), ABCC1 (multidrug resistance related protein 1, MRP1) und dem Cholesterintransporter ABCA1 in die Pathogenese der Alzheimer Krankheit involviert, da diese Proteine zur Clearance von Amyloid-ß aus dem Gehirn beitragen. Bekannt sind Zwischenfälle aus der Tiermedizin: Collies, Shelties und Bobtails dürfen nicht mit dem Antiparasitikum Ivermectin behandelt werden, da bei diesen Hunderassen häufig Gendefekte des ABCB1-Gens beobachtet werden. Das neurotoxische Ivermectin ist ein p-Glykoproteinsubstrat [6], kann aufgrund fehlender P-Glykoproteinfunktion bei den Hunden die Blut-Hirn-Schranke durchqueren und zum Tod der behandelten Tiere führen. Aufgrund seiner außerordentlich wichtigen Funktion bei der körpereigenen Abwehr toxischer Substanzen und der Entwicklung von Resistenzen bei Chemotherapie galt P-Glykoprotein in den letzten Jahren als Target beim Versuch, durch seine Blockade auch klinisch eine Zytostatikaresistenz aufzuheben, allerdings mit bislang eher enttäuschenden Ergebnissen. Ein interessantes Phänomen wird bei der Behandlung von Epilepsie beobachtet. Hier kommt es im Laufe der Behandlung mit Antiepileptika verstärkt zu Therapieresistenzen, vermutlich durch eine Induktion von ABC-Exportproteinen im Bereich epileptischer Foci, wodurch die Konzentration der Antiepileptika herabgesetzt wird [7]. An diesen wenigen Beispielen wird deutlich, welch entscheidende Rolle Transportproteine für Physiologie, Pathophysiologie, Toxikologie oder Arzneimittelwirkung spielen. Unser Wissen um diese Proteine hat in den letzten beiden Dekaden immens zugenommen, dennoch sind noch viele Fragen offen – wie funktioniert z.?B. ihre Substraterkennung, über welche Signalkaskaden werden sie reguliert, wie lassen sich Erkenntnisse zur Struktur dieser Proteine für ein rationales Drug Design nutzen? Die Beantwortung dieser Fragen wird helfen, ihre physiologische Rolle besser zu verstehen, Pathomechanismen von Krankheiten aufzuklären und neue Strategien der Wirkstofffindung zu entwickeln.
Literatur
Foto: |
L&M 3 / 2014
Das komplette Heft zum kostenlosen Download finden Sie hier: zum Download Die Autoren:Weitere Artikel online lesen

NewsSchnell und einfach die passende Trennsäule finden
© Text und Bild: Altmann Analytik ZEISS stellt neue Stereomikroskope vor
ZEISS stellt zwei neue kompakte Greenough-Stereomikroskope für Ausbildung, Laborroutine und industrielle Inspektion vor: ZEISS Stemi 305 und ZEISS Stemi 508. Anwender sehen ihre Proben farbig, dreidimensional, kontrastreich sowie frei von Verzerrungen oder Farbsäumen. © Text und Bild: Carl Zeiss Microscopy GmbH |








