

|
Bio&Biotech
 >
Versteinertes Leben
>
Versteinertes Leben
Versteinertes LebenMit biogenen Mineralen auf ZeitreiseAls ich während meines Mineralogiestudiums das erste Mal das Wort „biogene Minerale“ gehört habe, war ich der Meinung, dass diese beiden Worte selbst erklärend genug sind, um genau zu beschreiben, was gemeint ist. Wenn dazu dann noch als Beispiele z.B. Muschelschalen oder Korallen genannt werden, erst recht. Ein Mineral ist eben eine kristalline chemische Verbindung und die Biologie ist die Wissenschaft des Lebendigen – und diese Materialien haben eben mit beidem etwas zu tun, ganz einfach. Dass diese beiden Worte jedoch nicht annähernd die Komplexität dieser Materialien erahnen lassen, wurde mir aber in dem Moment klar, als ich vor über 15 Jahren damit angefangen habe, mich mit diesen Materialien genauer auseinanderzusetzen. Bevor wir den strukturellen Aufbau biogener mariner Karbonate etwas genauer betrachten, zunächst ein paar Worte dazu, was diese für viele Forschungsgebiete so interessant macht. Biogene Minerale als Helfer bei der Klimarekonstruktion 
Biogene marine Karbonate, gebildet von Organismen wie Muscheln, Schnecken, Korallen oder auch sehr kleinem, nur ein paar Mikrometer großem Plankton (wie etwa Foraminiferen oder Coccolithophoriden), kommen nicht nur in großen Teilen der Meere vor, sondern auch abgelagert in deren Sedimenten. Genau diese in den Sedimenten begrabenen biogenen Karbonate stellen eines der wichtigsten Fenster in die Vergangenheit unseres Planeten dar. Denn auch wenn diese biogenen Karbonate hauptsächlich aus Kalziumkarbonat (CaCO3) bestehen, so bauen sie in Spuren auch viele andere Elemente ein, die im Meerwasser vorhanden sind. Dieser Einbau geschieht jedoch nicht völlig willkürlich, sondern es zeigt sich bei näherem Hinsehen, dass oft ein systematischer Zusammenhang zwischen dem Einbau und den Umweltbedingungen, die zu Lebzeiten des kalzifizierenden Organismus herrschen oder eben geherrscht haben, besteht. So ist z.B. der Einbau von Magnesium in die Kalkschale von vielen Foraminiferen abhängig von der Wassertemperatur. Damit erhalten wir durch den Magnesiumgehalt, den wir in diesen biogenen Karbonaten messen können, ein perfektes Paleothermometer. Mit anderen Worten: die Möglichkeit, Wassertemperaturen an bestimmten Stellen zu bestimmten Zeitpunkten in der Vergangenheit zu bestimmen. Hierbei stellt der Magnesiumeinbau zur Temperaturbestimmung nur einen von vielen so genannten Paleoproxies dar, die zur Paleoklimarekonstruktion herangezogen werden. Viele weitere Elemente oder Isotope können Aufschluss über Umweltparameter wie z.B. den Salzgehalt oder Nährstoffgehalt zum Bildungszeitpunkt der biogene Karbonate geben, und liefern uns somit die grundlegenden Informationen, die wir benötigen, um die Funktionsweise der Geosphäre zu verstehen. Letzteres ist eine Grundvoraussetzung, um die durch den Menschen hervorgerufenen Veränderungen für die Zukunft abschätzen zu können. Auch wenn das Grundprinzip, wie Paleoproxies funktioniert, recht einfach klingt, stellt es für die Forscher eine große Herausforderung dar, daraus die richtigen Daten abzuleiten. Ein Grund dafür ist, dass der Zusammenhang zwischen einem Umweltparameter (nehmen wir als Beispiel wieder die Wassertemperatur) und dem Elementeinbau (also hier das Magnesium) für jede Art anders gestaltet ist und oft auch nicht ausgeschlossen werden kann, dass noch andere als der gesuchte Umweltparameter Einfluss auf den Einbau haben können. In der Praxis müssen daher aufwändige, so genannte Kalibrierungsexperimente, gemacht werden, bei denen z.B. eine bestimmte Foraminiferenart bei verschiedenen Temperaturen gezüchtet und das genaue Verhältnis zwischen Temperatur und Magnesiumeinbau für diese Art bestimmt wird. Weitere Experimente müssen dann im Anschluss zeigen, ob auch andere Parameter Einfluss auf den Einbau eines bestimmten Elements haben. Man kann sich leicht vorstellen, dass dies nicht für alle Arten und Parameter möglich ist. Nicht zuletzt deshalb, da es für viele Arten nicht möglich ist, diese unter Laborbedingungen zu halten. Zum Beispiel läuft bei manchen Arten die Bildung des biogenen Karbonats viel zu langsam ab, um dies in einem normalen Laborexperiment zu untersuchen oder der Organismus wächst außerhalb seines natürlichen Habitats überhaupt nicht. Biogene Minerale als mögliche Opfer der Ozeanversauerung Seit wenigen Jahren sind biogene marine Karbonate aber auch in den Fokus eines weiteren Forschungsgebietes gerückt, der Ozeanversauerung. Durch den Anstieg der CO2-Emission, der auf anthropogene Aktivitäten zurückgeht, löst sich auch mehr CO2 in den Meeren, was wiederum zu einer Erhöhung ihrer Kohlesäurekonzentration und damit einem Abfall des pH-Wertes führt. Bei einem weiteren CO2-Anstieg, momentan ist etwas anderes nicht zu erkennen, ist auch ein Einfluss auf kalzifizierende Organismen zu erwarten, da die Löslichkeit für Karbonate steigt. Hier stellt sich nun die Frage, inwieweit auch die Bildung der biogenen marinen Karbonate davon betroffen ist. Viele der wissenschaftlichen Fragen, welche durch diese unterschiedlichen Forschungsrichtungen hervorgebracht werden, erfordern ein sehr detailliertes Wissen über den strukturellen Aufbau mariner biogener Karbonate. Biogene Minerale einfach nur als Minerale zu sehen, die eben unter dem Einfluss biologischer Prozesse gebildet wurden, reicht dabei bei Weitem nicht aus. Was ist also so ein biogenes Mineral genau? Polymorphe, die vielen Variationen des Kalziumkarbonat Ich beziehe mich nachfolgend auf das Kalziumkarbonat. Es gibt zwar noch Karbonate anderer Elemente, jedoch stellt das Kalziumkarbonat das am weitesten verbreitete Karbonat dar. Karbonate sind chemisch gesehen CaCO3, bestehen also aus den Elementen Kalzium (Ca), Kohlenstoff (C) und Sauerstoff (O). Es gibt jedoch verschiedene Polymorphe, die sich in ihrer chemischen Zusammensetzung nicht unterscheiden, aber unterschiedliche Kristallstrukturen haben. Kalziumkarbonat kann in Form von sechs verschiedenen Polymorphen vorkommen. Hier sollen aber nur drei erwähnt werden, da diese später noch eine Rolle spielen werden: Vaterit, Aragonit und Kalzit. Man sagt zwar oft, dass sich Polymorphe „nur“ in der Kristallstruktur unterscheiden, dieser Unterschied hat jedoch weit reichende Konsequenzen für die chemischen und physikalischen Eigenschaften der Minerale. So unterscheidet sich sowohl die Löslichkeit der Polymorphe als auch die Möglichkeit, andere Elemente (in kleinen Mengen) an Stelle des Kalziums einzubauen, erheblich. Es sei noch erwähnt, dass die ursprüngliche Definition für Mineral sich nur auf natürlich (durch geologische Vorgänge) gebildete, kristalline chemische Verbindungen bezogen hat, inzwischen aber (nach einigen Diskussionen) auch von biogenen Mineralen gesprochen wird. Es mag etwas kleinkariert klingen, dieses so herauszustellen, aber es steht exemplarisch dafür, wie sich unser Wissen über biogene Minerale entwickelt hat. Am Anfang ging man davon aus, dass die einzige Funktion des Organismus als biologischer Anteil darin besteht, einen Ort für die Karbonatbildung zur Verfügung zu stellen – als würde man einen Schluck Meerwasser nehmen und darin würde dann anorganisch das Karbonat ausgefällt, nur eben dass das Becherglas ein Kompartiment irgendwo in einem Organismus ist. Für diese erste Hypothese sprach, dass das Meerwasser von der Zusammensetzung her eigentlich für Kalzit übersättigt ist und aus rein thermodynamischen Überlegungen heraus Kalzit gebildet werden könnte. Der Grund dafür, dass in unseren Meeren nicht riesige Mengen Kalzit anorganisch ausgefällt werden, liegt unter anderem an dem hohen Magnesium- und Phosphatgehalt des Meerwassers, der die Kalzitfällung unterbindet. Lassen wir aber diesen Umstand zunächst einmal beiseite. Wenn also die Biomineralisation nichts anderes ist als das Aufnehmen von Meerwasser, um daraus Kalzit zu bilden, sollte der gemessene Einbau von anderen Elementen (die an Stelle des Kalziums eingebaut wurden) in der Tat vergleichbar sein mit der Fällung im Becherglas. Um es kurz zu mache: Es stellte sich heraus, dass dies nicht der Fall ist. Die gemessenen Werte unterschieden sich grundlegend. Dies ist auch der Grund für die Notwendigkeit der oben erwähnten Proxiekalibrierung, die gemacht werden muss, da der Elementeinbau artspezifisch ist. Aus dieser Erkenntnis heraus wurde der Begriff des vitalen Effektes („vital effect“) geprägt [1], der aussagt, dass der Elementeinbau (oder auch die Isotopenfraktionierung), die in einem biogenen Mineral gemessen wird, durch den Organismus beeinflusst wird. Wie weit geht aber nun dieser Einfluss? Lange glaubte man, dass es nur darum geht, die Zusammensetzung des Meerwassers so weit zu verändern (man denke nur an die erwähnte „zu hohe“ Magnesiumkonzentration), dass die Kalzitbildung stattfinden kann. Frühere Beobachtungen an biogenen Karbonaten haben diese Sichtweise auch nahegelegt. Eine klassische, immer noch sehr aussagekräftige Methode in der Mineralogie ist die Polarisationsmikroskopie. Hierzu wird die zu untersuchende Probe (z.B. ein Stück Gestein, welches ja aus unterschiedlichen Mineralen aufgebaut ist) auf einen Glasträger aufgeklebt und auf eine Dicke von nur ca. 40 µm geschliffen. Wenn die Mineralkörner so dünn geschliffen sind, kann man unter dem Mikroskop durch sie hindurchsehen. Wird dabei polarisiertes Licht (Licht, das nur eine Schwingungsebene hat) verwendet, erscheinen sie auf Grund ihrer optischen Eigenschaften farbig und man kann unter anderem die kristallografische Orientierung der Kristalle erkennen. Polarisationsmikroskopische Untersuchungen an Dünnschliffen von biogenen Karbonaten zeigen oft, dass große Bereiche dieselbe „Farbe“ und somit kristallografische Orientierung aufweisen, was typisch für Einkristalle ist. Diese Beobachtungen unterstützten die Annahme, dass biogene Kristalle wie bei einer anorganischen Fällung im Becherglas wachsen. Als man jedoch auf die geniale Idee kam, Bereiche, die man für einen einzelnen großen Kristall gehalten hat, zu ätzen (mit einer speziellen Lösung, die Karbonat löst, aber organische Verbindungen stabilisiert), wurden zur großen Überraschung Strukturen erkennbar die nicht ganz in das bisherige Bild passten. Lauter kleine parallele Bänder wurden sichtbar, die aus organischen Verbindungen bestehen mussten [2], [3]. Ermöglicht durch die Entwicklung analytischer Methoden mit immer höherer Ortsauflösung wie zum Beispiel der Rasterkraftmikroskopie (welche die Untersuchung von Strukturen mit wenigen Nanometern Größe möglich macht), zeigte sich, dass viele biogene Mineralien aus Kristallen mit einer Größe von nur wenigen hundert Nanometern aufgebaut sind. Diese kleinen Kristallite sind bei vielen Arten von einer hauchdünnen organischen Schicht umhüllt, bilden aber in ihrer Gesamtheit Strukturen, die mehrere Millimeter oder gar Zentimeter groß sind und uns makroskopisch als ein homogener anorganischer Kristall erscheinen. Dieser komplexe Aufbau biogener Minerale als „Verbundwerkstoff“ konnte bisher bei sehr vielen unterschiedlichen Arten wie Muscheln, Schnecken, Korallen oder Foraminiferen nachgewiesen werden. Es gibt doch mehr zu bedenken als gedacht Was hat das jetzt aber mit den Karbonaten als Proxiearchiv zu tun? Die schon eben erwähnte Entwicklung räumlich immer hochauflösender analytischer Methoden wie z. B. der Laser Ablations-Massenspektrometrie (hierbei wird mit einem Laserstrahl ein winziges, nur wenige Mikrometer großes Loch in eine Probe geschossen und der Staub aus diesem Loch dann mit einem Massenspektrometer untersucht), hat dafür gesorgt, dass man nicht nur, wie es in den Anfängen der Fall war, große Probenmengen untersuchen kann, sondern auch sehr kleine Probenbereiche.

Elektronenmikroskopische Aufnahme des angeschliffenen und geätzten Statolithen eines Riesenkalamars (Architeuthis). Diese Statolithen sind nur wenige Millimeter große biogene Minerale, die in den Lage- und Gleichgewichtsorganen vorkommen. Die zu erkennende Ringstruktur stellt Wachstumslinien dar, jedoch ist es bisher nicht gelungen zu bestimmen, welchen Zeitraum eine Wachstumslinie repräsentiert.
Dies hat für die Untersuchung von Proxyarchiven wie z.B. einer Muschelschale folgende Auswirkung. Ähnlich wie bei Baumringen kann man nicht nur einen Mittelwert (nehmen wir wieder unsere Temperatur) bestimmen, sondern kann sogar z.B. mit Jahresauflösung Temperaturänderungen zu Lebzeiten des Organismus bestimmen. Bei sehr langlebigen Archiven wie z.B. der Schale der Muschel Arctica islandica kann dies theoretisch einige hundert Jahre umfassen. Andererseits muss diese Inhomogenität der Probe berücksichtigt werden, um keine Fehler in der Interpretation der gemessenen Daten zu machen.

Abb. 1: Weiter oben habe ich schon einmal auf die drei Polymorphe Vaterit, Aragonit und Kalzit und deren Unterschiede im Elementeinbau hingewiesen. Dies geschah natürlich nicht ohne Grund. Es ist nun nicht so, dass biogene Karbonate nur aus Kalzit bestehen. Viele, wie z.B. Korallen und Muschelschalen, bestehen aus Aragonit, die Schalen vieler Arten sogar aus Kalzit und Aragonit oder, wie wir zeigen konnten, in einem Fall sogar aus Vaterit, Aragonit und Kalzit [4].
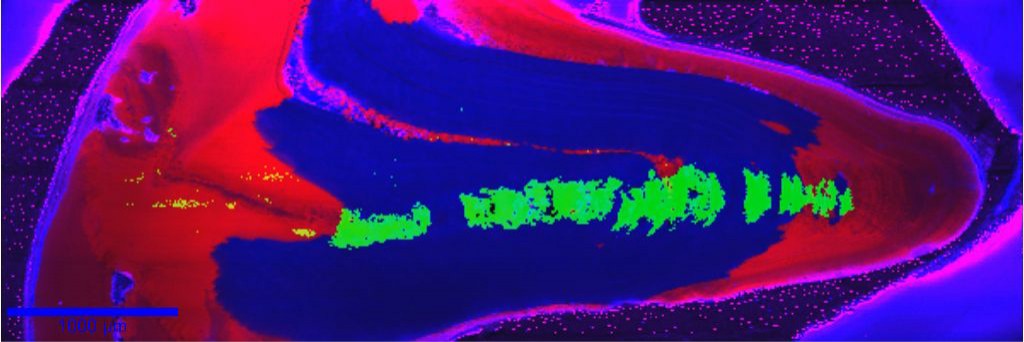
Abb. 1: Die Methode, die ich für eine der idealsten für die Untersuchung biogener Strukturen halte, ist die konfokale Raman-Mikroskopie. Diese Methode funktioniert in etwa so: Licht (normalerweise von einem Laser) trifft auf eine Probe und wird überwiegend reflektiert (dieser Teil interessiert uns eigentlich nicht), ein sehr kleiner Teil des Lichtes (Photonen) geht aber eine Wechselwirkung mit den Molekülen der Probe ein (regt bestimmte Molekülschwingungen an). Diese Wechselwirkung, der so genannte Raman-Effekt, kann nun genutzt werden, um Verbindungen zu identifizieren. Unter anderem kann man verschiedene Polymorphe unterscheiden und ihre Verteilung mit der sehr hohen örtlichen Auflösung von wenigen hundert Nanometern bestimmen. Dies ist jedoch nicht die einzige Information, die man bekommen kann. Man erhält gleichzeitig noch Informationen über die kristallografische Orientierung der Minerale [5] (wie mit der oben erwähnten Polarisationsmikroskopie, nur ohne Dünnschliffe anfertigen zu müssen [6], was bei manchen biogenen Mineralen recht schwierig sein kann). Zusätzlich kann aber auch die Verteilung mancher organischer Verbindungen visualisieren, sowie Informationen über die chemische Zusammensetzung der Verbindungen ableiten.

Abb. 1:

Abb. 1:

Abb. 1: Je mehr wir wissen, umso mehr neue Fragen ergeben sich
Das Bild, das wir hierdurch über den strukturellen Aufbau biogener Minerale erhalten, ist einzigartig und faszinierend. Jedoch ergeben sich aus jeder neuen Erkenntnis mindestens doppelt so viele neue Fragen. Fragen wie: Wodurch erlangt der Organismus so viel Kontrolle über die Bildung dieser biogenen Strukturen? Welche organischen Verbindungen sind dafür verantwortlich, welches Polymorph an genau welcher Stelle gebildet wird?
Literatur Foto: © masterfile.com | Little_Desire |
L&M 3 / 2013
Das komplette Heft zum kostenlosen Download finden Sie hier: zum Download Der Autor:NewsSchnell und einfach die passende Trennsäule finden
© Text und Bild: Altmann Analytik ZEISS stellt neue Stereomikroskope vor
ZEISS stellt zwei neue kompakte Greenough-Stereomikroskope für Ausbildung, Laborroutine und industrielle Inspektion vor: ZEISS Stemi 305 und ZEISS Stemi 508. Anwender sehen ihre Proben farbig, dreidimensional, kontrastreich sowie frei von Verzerrungen oder Farbsäumen. © Text und Bild: Carl Zeiss Microscopy GmbH |

